



.jpg)





ANOMALIES D’ORIGINE ANATOMIQUE 1ère partie
peut-être.
d’origine congénitale ou acquise. Cette dernière est beaucoup plus fréquente. Les
malformations anatomiques affectent généralement des individus et non des
groupes de vaches. Il est donc peu probable
qu’elles aient un effet réel sur la fécondité d’un troupeau.
1. ANOMALIES CONGENITALES
--
1.1.
Agénésie ovarienne (impuissance ; impossibilité d’engendrer).
Quoique
très rare, l’un ou les 2 ovaires peut-être.
atteints. Le tractus génital est infantile et la cyclicité est inexistante.
1.2.
Hypoplasie ovarienne
L’un
ou les 2 ovaires sont petits et lisses (OPL) afonctionnels. Conditionnée
par un gène autosomal récessif en association avec le ou les gênes responsables
de la décoloration du pelage, une forme d’hypoplasie ovarienne a été décrite
dans le bétail Highland (hautes terres) suédois. Cette anomalie affecte les ♀
et ♂ (13%). La principale anomalie se situe au niveau de l’épithélium
germinatif, il n’y a pas de développement d’ovogonies ni de follicules
primordiaux.
L’hypoplasie
ovarienne est uni ou bilatérale (9% des cas), l’ovaire gauche étant
inexplicablement plus fréquemment atteint que le droit. Quand les 2 ovaires
sont atteints, le tractus génital est infantile et le cycle sexuel est absent.
Les tétons sont petits et de consistance dure. Le bassin est étroit. La
confirmation du diagnostic requerra deux examens à un mois d’intervalle. La
réforme des individus atteints est conseillée.
L’hypop.
ovarienne doit ê différenciée de l’anoestrus vrai rencontré chez
certaines génisses. Dans l’anoestrus vrai, les ovaires ne sont pas aussi
petits, leur surface est lisse et non sous forme de petites masses fibreuses,
leur forme est circulaire ou ovalaire plutôt qu’allongée sous forme de masses
fibreuses. Le tractus génital tubulaire est plus développé.
En
cas d’anoestrus vrai, pour peu que les génisses soient en bon état physique, le
cycle sexuel se manifestera lorsque les conditions d’environnement sont
satisfaisantes. Des cas extrêmes d’hypoplasie gonadique se rencontrent chez le
bovin free-martin.
1.3.
Anomalies affectant les trompes
Les
oviductes semblent être les plus fréquemment touchés par les défauts d’origine
congénitale : l’aplasie unilatérale, la duplication des trompes, l’aplasie
segmentaire, l’obstruction bilatérale de l’oviducte, l’hydrosalpinx…
1.4.
anomalies affectant l’utérus et le col
- L’utérus didelphe
La
fusion incomplète de la portion postérieure des canaux de Müller peut entraîner
l’apparition d’un double vagin, d’un double col chacun s’ouvrant séparément
dans une corne utérine (utérus didelphys). Parfois, cette anomalie peut ne
concerner que le col.
La
duplication partielle intéressant l’orifice cervical externe est plus courante.
Elle serait d’origine héréditaire. Cette anomalie peut entraîner une dystocie
et une mortinatalité. Parfois les membranes placentaires restent accrochées
entre les 2 orifices.

Double
col plus bride (Hanzen, 2004)
·
Utérus unicorne
L’utérus
unicorne a été décrit. Cette anomalie peut s’accompagner d’un état d’anoestrus
si l’ovulation et le corps jaune ont lieu du côté ipsilatéral à la corne
absente. Cette anomalie n’empêche pas la gestation. Une solution consiste à
pratiquer l’ovariectomie unilatérale.

(Hanzen,
2004).
1.5.
La maladie des génisses blanches
Dénommée
par les Anglo-saxons, White Heifer Disease (WHD).
Des défauts de développement des canaux de Müller (paramesonephrotique) sont à
l’origine de différentes anomalies de l’oviducte, l’utérus, le cervix et le
vagin.
Cette
pathologie congénitale est liée à un gène autosome récessif dont l’action
serait associée ou favorisée par la présence du facteur blanc Shorthorn.
Les
anomalies vaginales sont de
loin les plus fréquentes. Le vagin peut être absent, raccourci ou interrompu.
La jonction vulvo-vaginale est souvent interrompue par une cloison appelée
hymen, non perforée ou percée d’un ou de deux orifices. Dans certains cas,
cette interruption se limite à une ou deux brides. La persistance de l’hymen peut conduire à
l’accumulation des sécrétions utérines en arrière du col. Il s’ensuit une
distension vaginale (mucocolpos)
responsable d’efforts expulsifs et de fièvre, pouvant dans les cas les plus
graves empêcher une miction ou une défécation normale. Le liquide aseptique peut se contaminer lors
de saillies malencontreuses. Ce type de lésion vaginale a également été décrit
en de très rares occasions dans l’espèce équine.

Mucopolpos
(distension vaginale), in : Hanzen, 2004.
Les
anomalies utérines sont plus
rares et consistent en une aplasie segmentaire localisée ou non, plus ou moins
étendue et responsable dans certains cas d’une dilatation kystique de
la corne utérine (hydromètre). Si le phénomène est unilatéral, la corne
droite est plus souvent modifiée que la gauche. La corne saine peut être
gestante. Le corps, col et vagin sont parfois totalement absents. Parfois on peut découvrir dans la corne un histerolithe résultant de
la dessiccation du mucus accumulé dans la corne. L’aplasie segmentaire d’une corne a été
décrite chez la jument. Le cas était associé à une tétralogie de Fallot.
Le
diagnostic d’aplasie segmentaire d’une corne utérine peut être posé par
palpation ou plus spécifiquement par instillation de liquide (sérum
physiologique) dans la corne pour confirmer l’absence de dilatation de cette
corne.

Bride
(Hanzen, 2004).
Les
ovaires conservent leur fonctionnalité. L’ovaire ipsilatéral à la corne
aplasique peut être porteur d’un corps jaune. Il en résulte un anoestrus
permanent du fait de l’absence de sécrétions de prostaglandines ou en aplasie
segmentaire. Etant donné l’absence d’anomalies ovariennes, les animaux atteints
présentent le plus souvent des chaleurs normales ce qui permet de les
différencier aisément des free-martin.
1.6.
Les pathologies de la différentiation sexuelle
Certains
animaux sont dits intersexués car
ils présentent en même temps des caractéristiques mâle et femelle. De telles
anomalies sont d’ordre génétique ou chromosomique ou imputable à une exposition
hormonale anormale lors de la différenciation sexuelle. Le terme hermaphrodite est plus spécifiquement
réservé aux animaux dotés des deux sexes anatomiques et fonctionnels.
Un bref rappel physiologique s’impose. Le déterminisme du
sexe est lié à la présence sur le chromosome Y d’un gène
appelé SRY (Sex determining region of Y) ou encore TDF (Testis Determining
Factor) et sur le bras court du chromosome X d’un gène appelé DSS (Dosage
Sensitive Sex Reversal). Le premier est un gène de masculinisation :
il induit une différenciation des ébauches génitales vers le type mâle. Le
second est un gène de féminisation : il induit la différenciation
des ébauches génitales vers un type femelle. Cette différenciation femelle
implique l’absence du gène SRY mais surtout la présence du gène DSS. Il est
important de préciser également que le gène SRY ne peut réprimer qu’un seul
exemplaire du gène DSS. Dès lors si à la suite d’une aberration génique sur le
chromosome, ce gène existe en deux exemplaires chez un individu de type XY, il
peut en résulter une différenciation incomplète du type mâle et la présence
d’anomalies histologiques au niveau des gonades, pouvant dès lors présenter du
tissu testiculaire et/ou ovarien. Dans toutes les espèces de mammifères, le
sexe mâle se différencie plus tôt que le sexe femelle. Chez le mâle, les
conduits et glandes annexes proviennent des canaux de
Wolff (canaux mesonephrotiques) tandis que
chez la femelle, l’oviducte, l’utérus et le vagin dérivent des canaux de
Müller (conduits paramesonephrotiques). Les canaux de Wolff sont donc de
simples vestiges chez la femelle (canaux de Gaertner au niveau du
plancher vaginal). Dans l’un et l’autre sexe, le système génital externe
provient du tubercule génital qui donnera le pénis chez le mâle et le clitoris
chez la femelle et des bourrelets génitaux qui constitueront le fourreau et le
scrotum chez le mâle et les lèvres vulvaires chez la femelle.
Chez le fœtus mâle, le gène SRY induit la synthèse d’une protéine
responsable de la différenciation de la gonade indifférenciée en testicule et
du développement des cellules de Sertoli et de Leydig. Les premières
synthétisent l’hormone MIS (Müllerian inhibiting substance) encore appelée AMH
(Anti Mullerian Hormone). Les secondes synthétisent les androgènes responsables
de la poursuite du développement des canaux de Wolff. Chez le foetus femelle,
en l’absence du TDF on observe une différenciation de la gonade primitive en
ovaire. Etant donné, l’absence de
sécrétion de MIS, on assiste au développement des canaux de Müller tandis que les
canaux de Wolff régressent étant donné l’absence de testostérone (Cours Hanzen,
OGA 2004).
1.6.1.
Intersexualité
Les
animaux intersexués sont classés en hermaphrodites
vrais et en pseudo-hermaphrodites.
Les premiers se caractérisent par la présence chez un même individu de gonades
des deux sexes, isolées ou associées en une glande unique : l’ovotestis.
Les seconds se caractérisent par la présence de gonades d’un sexe et de voies
génitales ou d’organes génitaux des deux sexes ou du sexe opposé. On parle de pseudo-hermaphrodisme mâle quand les
glandes sexuelles sont des testicules et de pseudo-hermaphrodisme femelle si les gonades sont des ovaires.
L’hermaphrodisme
vrai ambiglandulaire (alterne si testicule d’un coté et ovaire de l’autre, uni
ou bilatéral selon que l’on a à la fois testicule et ovaire d’un seul ou des
deux côtés) ne se rencontre pratiquement que chez le porc. Il serait imputable
à un gène récessif. Le plus souvent les vrais hermaphrodites ont une apparence
externe de type femelle.
Le
pseudo-hermaphrodisme masculin est le plus fréquent, surtout chez la chèvre
(associé au caractère sans cornes) et le porc.
Il a été décrit chez le cheval. Les gonades sont des testicules tandis
que les voies génitales internes et externes, le plus souvent hypoplasiques
sont de type femelle. Parfois dans l’espèce bovine on peut observer une
fermeture incomplète du canal de l’urètre au niveau périnéal ou scrotal (hypospadias). Il s’agirait d’une
insuffisance de réponse de l’urètre à l’influence de la dihydrotestostérone.
90
à 95 % des veaux femelles co-jumelles d’un veau mâle présentent une
masculinisation de leur tractus génital et sont stériles. Cette pathologie est
pratiquement spécifique de l’espèce bovine. Son appellation dériverait du
dialecte écossais. Le terme « free » signifie stérile et le terme
« martin » se rapporte sans doute au fait que les animaux stériles
étaient le plus souvent abattus à la fête de la Saint Martin (11 novembre).
L’animal femelle concerné présente
les caractéristiques suivantes : (1) Les gonades sont de volume réduit et
ne présentent aucune structure ovarienne. La glande peut être une structure
testiculaire mais la spermatogenèse en est absente. (2) Les structures
provenant des canaux de Müller sont sous-développées et celles dérivant des canaux
de Wolff sont anormales. (3) Les organes génitaux externes sont de type femelle
mais le périnée est allongé, la vulve petite et garnie de poils à la commissure
inférieure, le clitoris plus ou moins hypertrophié, saillant et renfermant
parfois l’urètre. La glande mammaire est atrophiée. La conformation extérieure
de l’animal rappelle celle du castrat (tête forte, cornes et encolure
développée).
Le
diagnostic peut être posé au cours du 1er mois suivant la naissance
au moyen d’un fin spéculum vaginal voire plus simplement avec un tube à prise
de sang (Test tube). Chez le veau normal, âgé de 12 à 28 jours, le vagin a une
longueur de 12 à 15 cm
(5 à 6 pouces )
alors que le vagin de la free-martin est borgne et que sa longueur est de 4 à 5 cm (2 à 3 pouces ).
L’apparition
du free-martinisme résulte de la présence de la formation d’anastomoses
vasculaires entre les placentas des deux foetus. Il en résulte des échanges
cellulaires et hormonaux entre le foetus mâle et femelle. En effet il a été
démontré que le free-martin présente deux populations cellulaires distinctes
l’une de type XX et l’autre de type XY en provenance du cojumeau mâle. Cette
présence serait davantage le témoin de l’anomalie que son facteur étiologique.
En fait, il semble bien que le free-martinisme soit induit par le passage chez
le foetus femelle du facteur MIS (régression des canaux de Muller pendant la
phase d’inhibition qui débute vers le 50ème jour de gestation) et de
la DHT (hormone responsable de la phase de masculinisation qui débute vers le
75ème jour de gestation).
Le
pourcentage de chimérisme (lymphocytes) chez les veaux mâles co-jumeaux de veaux femelles est compris entre 5 et 95
%. Ces animaux présentent une fréquence beaucoup plus élevée d’anomalies de la
fonction testiculaire : réduction de la production spermatique,
augmentation de la fréquence des formes anormales, réduction de la motilité et
de la concentration.
Malgré
la présence d’anastomoses vasculaires entre les jumeaux et l’apparition d’un
chimérisme (présence de populations cellulaires provenant de deux organismes
différents), le free-martinisme n’a pas été décrit dans l’espèce équine.
Certains cas ont été signalés dans les espèces ovine et caprine et suspectés
dans l’espèce porcine.
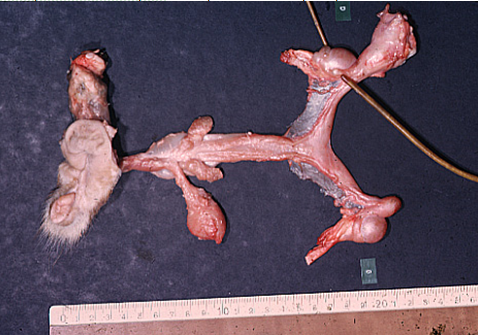
Freemartinisme
(Hanzen, 2004).

Freemartinisme (Hanzen,
2004).
1.6.3.
Hybrides interspécifiques
Le
nombre de chromosomes est spécifique de l’espèce. Pour rappel, le nombre de
chromosomes dans les espèces porcine, humaine, ovine, caprine et bovine, asine,
équine est respectivement de 38, 46, 54, 60, 62 et 64. Le mulet a 63
chromosomes.
Les
animaux hybrides sont dits eugénésiques
ou paragénésiques suivant que les deux
sexes ou que l’un des deux est fécond. Ils sont dits agénésiques lorsqu’ils sont totalement stériles. Rappelons que
les mulets et les bardots sont des hybrides de l’âne et du cheval, le mulet
provenant du croisement d’une jument et d’un âne mâle (baudet) et le bardot
d’un étalon et d’une ânesse. Les hybrides mâles sont stériles. Mulets et
bardots sont stériles pour les mâles en principe. On a signalé des cas de mules
qui se seraient reproduites avec succès avec des étalons.
Le
croisement d’un bélier avec une chèvre est possible mais le plus souvent la
gestation s’interrompt au bout de 6 semaines sous l’action d’anticorps
maternels. De même, le croisement du porc domestique et du sanglier donne
naissance à des hybrides fertiles.
Il
faut également signaler dans l’espèce caprine une particularité anatomique
donnant à penser à l’intersexualité. Il s’agit de la gynécomastie ou milking buck syndrome. Le bouc présente un
développement excessif des bourgeons mammaires. Ces animaux sont fertiles et
transmettent l’anomalie à leur descendance. L’affection a été décrite dans la
race Nubienne.

Gynécomastie chez le
bouc (Hanzen, 2004).
1.7.
Kystes paraovariens
Divers
kystes paraovariens d’origine congénitale, reliquats des conduits
mésonéphrotiques ont été décrits chez la truie, la vache, la jument. Leur
taille est comprise entre quelques millimètres à plusieurs centimètres.
L’aplasie bilatérale de l’infundibulum (portion ovarienne de l’oviducte) a été
décrite chez la vache. Il en résulte une accumulation de liquide dans la partie
distale de l’oviducte et de l’imperméabilité tubaire. De telles accumulations liquidiennes
(hydrosalpinx) peuvent également s’observer après une inflammation (métrites)
ou la formation d’adhérences autour de l’ovaire (césariennes).

1.8.
Atrésie vulvaire
peut-être
rencontrée chez les génisses de la race Holstein et Jersey. Elle est souvent
cause de dystocies ce qui nécessite césarienne / épisiotomie pour libérer le
veau. Une origine héréditaire n’est pas à écarter.

0 التعليقات:
Enregistrer un commentaire